Research
Mechanoelectrical transduction in hair cells
Electromechanical transduction in outer hair cells
Mechanoelectrical transduction in hair cells
Sound induced fluid vibrations in the cochlea are transduced into an electrical current by mechanosensitive ion channels located in the stereocilia. These channels are “gated” – opened and closed – as a result of deflection of the stereocilia. The mechanism for gating these channels is still unknown. Gating is purely mechanical. This is simply because of the requirement of extraordinarily high speed – 200 kHz in some bats and a “modest” 20 kHz in humans. Gating has been shown to be associated with extremely fine extracellular filaments, called tip links; their diameter is about 6 nm. Destruction of tip links, by for example sound overstimulation, leads to deafness.
We investigate mechanoelectrical transduction processes, employing the whole-cell patch-clamp technique in combination with fluorescent imagining of calcium uptake using a two-photon confocal laser-scanning microscope. We have shown that upon enzymatic or non-enzymatic destruction of the tip links, the channels appear to open, yielding a “tonic” current (Meyer et al., 1998). Recently, we showed that this tonic current is mechanosensitive and that the gating process is calcium dependent (Meyer et al., 2005). The data present evidence for an intracellular, calcium-dependent elastic gating element.
Electromechanical transduction in outer hair cells
A change of membrane potential of the outer hair cell (OHC) is transduced into a mechanical force, causing the cell body to contract and elongate. This electrically induced motion, called somatic electromotility, is believed by many researchers to be the basis of the exquisite frequency selectivity, sensitivity and dynamic range of the cochlea. The electromechanical force produced by the cell is thought to counteract damping forces in the cochlea, such as those inherent to the motion of fluids and cells, to produce the extraordinary mechanical tuning of the cochlea. The motor molecule responsible for somatic electromotility was cloned by the team of Peter Dallos; it belongs to a gene family, the solute carrier family 26, which encodes anion transporters and related proteins; it is called prestin. There are at least 6,000 of these molecules per square micrometer of cell surface. It is not known how the motion of all these molecules is coupled to the cytoskeleton, to generate the somatic electromotility. As with mechanoelectrical transduction in the stereocilia, the process must be exceedingly fast because it is required to work up to ultrasonic frequencies. Destruction of this transducer leads to deafness.
We investigate the electromechanical transduction process und employ laser Doppler vibrometry together with atomic-force spectroscopy. Using the vibrometer we showed that somatic electromotility can follow changes of the membrane potential cycle-for-cycle up to at least 100 kHz (Frank et al., 1999). We thereby established that the soma of the OHC has the necessary molecular machinery to generate electromechanical responses – with little attenuation and time delay – well-above the frequency limit of hearing. Since then we have developed an innovative method to measure the mechanical properties of biological structures up to at least 40 kHz (Scherer et al., 2000; Scherer and Gummer, 2004a). This technique employs an atomic force cantilever and has general applications outside the area of hearing research; it is particularly attractive for investigation of biological structures in a fluid. We are applying the technique to investigate mechanical and cell biological properties of cellular and membranous structures in the cochlea.
The requirements of high sensitivity, frequency selectivity and dynamic range can only be achieved by means of a mechanical amplifier in the cochlea. There are two schools of thought as to how the amplification might be achieved. Both hypotheses rely on the generation of mechanical force from an electromechanical transducer in the outer hair cells: one hypothesis maintains that the transducer is located in the soma and the other asserts that it is in the stereocilia. The soma hypothesis has the advantage that the outer hair cells are known to be electromotile over the entire functionally relevant frequency range (Frank et al., 1999), but the disadvantage that the receptor potential – the drive for the somatic electromotility – is low-pass filtered by the electrical impedance of the basolateral cell membrane (Preyer et al., 1996). Low-pass filtering, relative to stereocilia displacement, attenuates and delays the receptor potential to such an extent that somatic electromotility alone can not be responsible for cochlear amplification. The hypothesis that the relevant high-frequency electromechanical transducer is located in the stereocilia has the advantage that it does not suffer from low-pass filtering by the stereocilia membrane. However, it has the disadvantage that, to date, there is no direct experimental evidence that the stereocilia are electromotile at high frequencies. This having been said, there is evidence from the group of Robert Fettiplace that the stereocilia of outer hair cells possess electromechanical transducers. Whether these transducers can operate at sufficiently high frequencies and can inject enough mechanical force into the organ of Corti remains to be discovered.
We investigate mechanisms of cochlear amplification by the outer hair cells. Based on three-dimensional vibration measurements, we reported that the tectorial membrane presents resonant motion in the radial direction (Gummer et al., 1996; Hemmert et al., 2000a). We suggested that the temporal properties of this motion might compensate for the delay of the somatic electromechanical force produced by the electrical impedance of the basolateral membrane of the outer hair cell. These experiments gave the first hint as to how the electromechanical force might be coupled into the organ of Corti to yield cochlear amplification. Since damping and inertia of the organ of Corti might also cause severe attenuation and delay of the electromechanical force at high frequencies, we have also measured the mechanical impedance of the organ of Corti (Scherer and Gummer, 2004a). We found that for point loading, the organ of Corti moves like a purely viscoelastic material at all physiologically relevant stimulus frequencies; that is, there is no inertial component. Combining these impedance measurements with measurements of the velocity of the organ of Corti in response to intracochlear electrical stimulation, we recently found that the electromechanical force exerted at the reticular lamina has a broad resonant maximum (Scherer and Gummer, 2004b). We suggest that piezoelectric resonance in the outer hair cell, as proposed by Bill Brownell and colleagues, might be responsible for this force response. This resonance might compensate for the amplitude attenuation caused by the electrical impedance of the basolateral membrane of the outer hair cell. We are now investigating how the electromechanical force is coupled to the stereocilia of the inner hair cells.
Laseraudiometria
Desi este disponibil un numar mare de
teste audiometrice pentru
diagnosticul diferential al afectiunilor
auditive, nu este inca posibil sa localizam pozitia potentialelor
pierderi mecanice din
cohleea umana. Aceasta se datoreaza
faptului ca la om
urechea interna este inglobata in osul temporal
si din aceasta cauza nu este direct accesibila examinarii medicale.
Singurul indicator clinic al unei deficiente auditive prin lezarea
amplificatorului cohlear este pierderea
otoemisiunilor acustice.
Totusi, otoemisiunile acustice nu permit un
diagnostic diferential al amplificatorului cohlear – de exemplu: pentru a decide daca
defectul se datoreaza decuplarii
stereocililor
celulelor ciliate
externe din
membrana tectoria ce le acopera sau daca apare prin
disruptia complexului motor din peretele bazolateral al celulelor
ciliate externe, sau...
Ca punct de plecare pentru dezvoltarea unui diagnostic diferential al amplificatorului cohlear, am dezvoltat tehnica laserinterferometrica pentru a evalua impedanta mecanica de input a cohleei. Pentru a realiza acest lucru, efectuam masuratori ale vibratiei timpanului. Aparatul l-am denumit laseraudiometru. Principiul se bazeaza pe faptul ca impedanta de input a cohleei se comporta ca o impedanta ce incarca urechea medie, astfel incat perturbarile mecanice din cohlee teoretic ar trebui sa fie observabile in vibratia timpanului. Am reproiectat sistemul de masurare pe care l-am dezvoltat anterior (Rodriguez Jorge si colab., 1997), pentru a permite masuratori ale vibratiei timpanului aproape de pragul auditiv. Am observat prezenta unei componente cohleare in vibratia membranei timpanice. Experimentele continua cu asistenta nepretuita a pacientilor cu hipoacuzie neurosenzoriala.
Outer hair cells (OHCs), like many other polarized cell types, possess an endoplasmic network of tubules and vesicles. In general, it serves specialized functions, including protein synthesis, sequestration of calcium, production of steroids, storage and production of glycogen, and insertion of membrane proteins. This network appears to be more elaborate in OHCs than in other cells. The lateral cisternae are an integral part of the endoplasmic system of OHCs, and are involved in membrane turnover. Hair cells are known to have endocytic activity at both their apical (endocytosis) and synaptic (exocytosis) poles. The role of membrane recycling, and especially endocytosis, is not yet fully understood in OHCs.
We are investigating membrane turnover processes. To investigate vesicle transport in OHCs we use various fluorescent dyes; for example, FM1-43 as a membrane marker, DiOC6 for endoplasmic reticulum and neutral red as a lysosomal probe. Fluorescence is visualized with a two-photon laser-scanning microscope. FM1-43 reversibly stains the membrane; trapped vesicles can be visualized when the membrane is endocytosed. We have demonstrated endocytosis at the apical pole of OHCs; the destinations of endocytosed plasma membrane are the infracuticular zone, subsurface cisternae and the synaptic region (Meyer et al. 2001). The aim of present studies is to reveal the regulation of the intracellular traffic of trapped vesicles within OHCs.
Information from the experiments is being incorporated into models of sound processing in the cochlea, to understand function and develop new diagnostic tools.
A model of the organ of Corti is being developed together with Professor Charles Steele, University of Stanford. The model yields mechanical impedance data consistent with measured data (Scherer and Gummer, 2004a). Conversely, the model explains the experimental observation that the impedance is purely viscoelastic, without an inertial component. The model is being incorporated into a complete model of cochlear function.
At the diagnostic level, together with Professor Renato Nobili, University of Padua, we are investigating the origin of otoacoustic emissions. These are acoustic signals generated by the normal cochlea, either in the absence of acoustic stimulation (spontaneous emissions) or in response to acoustic stimulation (acoustically-evoked emissions) or electrical stimulation (electrically-evoked emissions). They are a by-product of the electromechanical action of the outer hair cells. Acoustically evoked otoacoustic emission testing gives an objective indication of whether the cochlear amplifier is functioning normally; it is particularly important for neonatal screening. The mechanisms generating the emissions are not fully understood, so that optimal stimulus paradigms are not yet available. This project is solving these problems.

Receptor current from an outer hair cell

Two-photon confocal
laser-scanning microscope
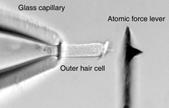
Force from
an outer
hair cell

Measurement of
mechanical impedance
Vibration of the organ of Corti
![]()
![]()

Laseraudiometer
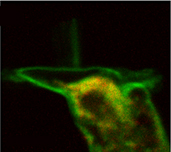
Double-labeling of an outer hair cell

3-D representation
of a hydrodynamic cochlea
model